Home » Wheat
Category Archives: Wheat

Banding P for Acidic Soils: Its not the time to be paying for poor practice.
I am bringing this topic back to the surface now with the current outlook on phosphorus fertilizer. If you have heard its not only becoming more expensive but the supply is short and will likely stay short through summer into the fall, which wont help prices. So this year’s wheat crop, we need to be prepared to be smart with Phosphorus, and applying an extra 30lbs to band aid for soil acidity should not be in the cards. Look at it this way, if the phosphorus was at $0.66 a lb that $20 that could be spent on a ton of lime. That lime will last 3-5 years, while that P needs to be added every year. Not only that, but the lime will help root growth (better when we dry up), produce significantly more biomass, and make the phosphorus you’ve applied in the past available again for plant uptake. So make the plans now to soil sample as soon as this crop is off, you can get a soil test recommendation and plan for the lime trucks. This is also not the year to just apply phosphorus for the sake of applying. Soil tests are inexpensive relative to buying excess fertilizer.
Current quotes on 4.24.26 are at $0.54 + per lbs P2O5 with DAP at $830 a ton.
Quick Fertilizer Price Calculation:
Urea at $860 a ton means N is $0.93 a lbs.
DAP at $830 has $334 worth of $0.93 nitrogen and $495 of phosphorus at $0.54 a lb.
Banding P as a band-aid for soil acidity, not so cheap now.
Original Blog Posted in 2021
Whoi Cho, PhD student Ag Economics advised by Dr. Wade Brorsen
Raedan Sharry, PhD Student Soil Science advised by Dr. Brian Arnall
Brian Arnall, Precision Nutrient Management Extension.
In 2014 I wrote the blog Banding P as a Band-Aid for low-pH soils. Banding phosphate to alleviate soil acidity has been a long practiced approach in the southern Great Plains. The blog that follows is a summary of a recent publication that re-evaluated this practices economic viability.
Many Oklahoma wheat fields are impacted by soil acidity and the associated aluminum (Al) toxicity that comes with the low soil pH. The increased availability of the toxic AL3+ leads to reduced grain and forage yields by impacting the ability of the plant to reach important nutrients and moisture by inhibiting root growth. Aluminum can also tie up phosphorus in the soil, further intensifying the negative effects of soil acidity. More on the causes and implication of soil acidity can be found in factsheet PSS-2239 or here (https://extension.okstate.edu/fact-sheets/cause-and-effects-of-soil-acidity.html). The acidification of many of Oklahoma’s fields has left producers with important choices on how to best manage their fields to maximize profit.

Two specific management strategies are widely utilized in Oklahoma to counter the negative impacts of soil acidification: Lime application and banding phosphorus (P) fertilizer with seed. While banding P with seed ties up Al allowing the crop to grow, this effect is only temporary, and application will be required every year. The effects of liming are longer lasting and corrects soil acidity instead of just relieving Al toxicity. Historically banding P has been a popular alternative to liming largely due to the much lower initial cost of application. However, as P fertilizers continue to increase in cost the choice between banding P and liming needed to be reconsidered.
A recent study by Cho et al.,2020 compared the profitability of liming versus banding P in a continuous wheat system considering the impacts that lime cost, wheat price and yield goal has on the comparison. This work compared the net present value (NPV) of lime and banded P. The study considered yield goal level (40 and 60 bu/ac) as well as the price of P2O5 fertilizer and Ag Lime. The price of P2O5 used in this study was $0.43 lb-1 while lime price was dictated by distance from quarry, close to quarry being approximately $43 ton-1 and far being $81 ton-1. For all intents and purposes these lime values are equivalent to total lime cost including application. Wheat prices utilized in the study were $5.10 bu-1 and $7.91 bu-1. It is important to note that baseline yield level was not considered sustainable under banded P management in this analysis. This resulted in a decrease in yield of approximately 3.2 bu ac-1 per year. This is attributable to the expected continued decline in pH when banding P is the management technique of choice.
The analysis in this work showed that lime application is cost prohibitive in the short term (1 year) when compared with banding P regardless of lime cost, yield goal level, and wheat value (within the scope of this study). This same result can be seen over a two-year span when yield is at the lower level (40 bu ac-1). While in the short-term banding P was shown to be a viable alternative to liming, as producers are able to control ground longer lime application becomes the more appealing option, especially when producers can plan for more than 3 years of future production. In fact, under no set of circumstances did banding P provide greater economic return than liming regardless of crop value, yield, or liming cost when more than 3 years of production were considered and only under one scenario did banded P provide a higher NPV in a 3-year planning horizon.
While historically banding P was a profitable alternative to lime application for many wheat producers the situation has likely drastically changed. At the time of writing this blog (09/17/2021) Diammonium Phosphate (DAP) at the Two Rivers Cooperative was priced at $0.78 lb-1. of P2O5. This is a drastic increase in P cost over the last year or so since Cho et al. was published in 2020. With P fertilizer prices remaining high it will be important for producers to continue to consider the value of liming compared to banded P. This is particularly crucial for those producers who can make plans over a longer time frame, especially those more than 3 years.
Addendum: As fertilizer prices have continued to rise a quick analysis utilizing the $0.78 lb-1 of P2O5was completed to consider the higher P fertilizer cost. Under this analysis an estimated decrease in NPV of approximately $38 an acre for P banding occurred. When considering this change in NPV, lime application becomes the more profitable option for alleviation of soil acidity symptoms even in the short term (assuming lime price values are equivalent to the previous analysis). This underlines the fact that it is imperative to consider the impact on profitability of the liming vs. banding P decision in the current economic climate for agricultural inputs.
Link to the Open Access Peer Reviewed publication “Banding of phosphorus as an alternative to lime for wheat in acid soil” https://doi.org/10.1002/agg2.20071
Protect Your Emerging Stands: True Armyworm Movement from Maturing Wheat to Summer Crops
Ashleigh M. Faris, Cropping Systems Extension Entomologist & IPM Coordinator
As the Oklahoma winter wheat crop reaches maturity, producers and crop consultants should prepare for the annual migration of true armyworm larvae. While true armyworms are a common fixture in small grains, their movement out of maturing wheat and into newly emerged corn, soybeans, and sorghum can lead to stand thinning or loss if not monitored closely.
True Armyworm Migration Timeline
True armyworm moths typically migrate into Oklahoma from the south in early spring with infestations typically occurring in late April through the first two weeks of May. The first generation is typically laid in winter wheat. Once the larvae currently finish their development in wheat, they will soon seek new food sources as the wheat crop dries down. This transition period is the most critical time for scouting summer crops, especially those adjacent to wheat fields.
True Armyworm Life Cycle and Identification
Armyworms overwinter as pupae or as mature larvae which pupate in the spring. Moths emerge in the spring, mate, and lay eggs in masses on hosts plants (mostly in the grass family). Female moths deposit their eggs in low-lying areas on wheat or pasture ground, as well as field margins or fields with dense, grassy weeds like Johnson grass. Larvae feed for about 4 weeks but do most of their damage during the last 10 days of this period. They then pupate in the soil. A new generation of moths emerges about 1 week later. There are 4 generations per year in Oklahoma.
True armyworms have a smooth body and can be variable in color, ranging from green, tan, orange, and black, with distinct pale orange or reddish stripes running along the sides (Figure 1). A key identifier is a dark diagonal band on each of the abdominal prolegs; there are four pairs of prolegs (Figure 2). The head capsule is light brown with a distinct “net-like” or honeycomb pattern of dark lines (Figure 2).

Figure 1. Four true armyworm larvae. One is dark (right) and three are light colored (left). Photo by Ashley Dean, Iowa State University Extension.

Figure 2. True armyworm. A) Dark band on prolegs. B) Orange head capsule with dark net-like pattern. Photos by Adam Varenhorst, Iowa State University Extension.
True Armyworm Management Cutoff in Wheat
A common question during this window is whether to treat armyworms in maturing wheat. Once wheat reaches the soft dough stage, the crop has generally accumulated its yield. Unless larvae are actively head-clipping (cutting the wheat heads off the stems), chemical control is rarely economical at this stage. Instead of treating the wheat, focus on young stands of summer crops. As wheat turns brown, larvae will move toward the nearest green tissue—often your emerging corn or sorghum.
Scouting, Damage, and Economic Thresholds for Summer Crops
Armyworms are whorl feeders in grass crops like corn and sorghum and will also feed on soybean leaves. True armyworms hide in the soil, crop residue, or whorls during the heat of the day and feed at in the early morning, evening or late when it is cool outside. When it is warm, larvae will hide in the soil, crop residue, or the whorl of corn plants. Large larvae consume more tissue but will generally be done feeding in a few days. Insecticides should target young, small larvae that will be feeding for a long time; however, you may see a range of larval sizes in a single field.
Corn, Sorghum, and Soybean Damage
True armyworm feeding typically begins at the leaf edges, leaving ragged holes and edges (Figure 3). As this leaf tissue is removed, the larvae will move to the upper leaves and continue feeding. True armyworms do not tunnel into the stalk and generally do not feed on the growing point of larger corn and sorghum plants. While not the preferred host, true armyworms will move into soybeans if no grasses are available. Larvae typically cause defoliation (Figure 4); however, soybeans are quite resilient to early-season leaf loss, but scout for stand-thinning if larvae are clipping seedlings.

Figure 3. True armyworm feeding on young corn plant. Photo by Adam Varenhorst, Iowa State University Extension.

Figure 4. Soybean leaves with true armyworm feeding damage. Photo by Meaghan Anderson, Iowa State University Extension.
Corn Threshold: Small plants typically recover from true armyworm feeding and outgrow the defoliation. Per Kansas State Extension, treatment is justified only when larvae are less than 1.25 inches long and present on 30% of plants with 5 – 6 extended leaves, or when 75% of plants have one or more larva per plant. There is risk of yield loss if defoliation during reproductive stages approaches the ear zone before hard dent. Lower thresholds may apply if the plants are subject to additional stresses.
Sorghum Threshold: Sorghum is very tolerant of defoliation, so insecticide control is rarely justified. For early infestations (5-7 leaf stage, prior to panicle development) at the vegetative stages where true armyworms may be in the whorl, do not initiate controls unless 40% or more of the plants in a field are infested. Because the worms are only defoliating at this point in the sorghum plant’s development, economic damage is not a concern and there would likely be no return on investment for spraying before panicle development.
Soybean Threshold: Once grasses are fed upon or harvested, true armyworms can turn tobroadleaf crops, including soybean. While soybean is not a preferred host, the growing point is exposed early in the season, making them susceptible to stand loss. Management is suggested if soybean defoliation is greater than 35% – 40% during the vegetative stages.
True Armyworm Insecticide Management Options for Summer Crops
True armyworm is generally easier to control with pyrethroids than fall armyworm. Ensure high-volume water (10-15 GPA ground) is used to get the product into the whorl or canopy where the larvae hide. Remember that most insecticides work via contact; if true armyworm larvae are feeding or hiding under dense residue, insecticides are unlikely to make contact and are ineffective. Target applications when larvae are actively feeding on foliage to ensure good contact. Follow all instructions on the insecticide label to ensure good control.
For a complete list of recommended insecticides and rates for these crops, please consult the following OSU Fact Sheets: CR-7167: Management of Insect and Mite Pests in Corn and Sorghum and CR-7115: Management of Insect and Mite Pests in Soybean.
The information given herein is for educational purposes only. Reference to commercial products or trade names is made with the understanding that no discrimination is intended and no endorsement by the Cooperative Extension Service is implied.
Mechanisms of Soil Fertility: Looking at Biologicals and MOA
Brian Arnall, Oklahoma State University Precision Nutrient Management
The use of biological products in commercial agriculture has expanded rapidly, with large corporations entering a space once dominated by smaller groups. This has created an arms race, with nearly every company offering a biological product. Over the past twenty years, I have had the opportunity to test products from the biggest groups with billions in backing, to solutions raised in stock tanks delivered in Braums milk jugs. It is critical to understand what is in the jug and the biological function it is expected to perform. Like herbicides, knowing the mode of action determines whether the product fits the intended purpose. No different than herbicides and knowing mode of actions. It’s important to know and understand that if you are trying to kill ryegrass 2.4-D, a broadleaf herbicide is not the right answer.
So what are we working with that’s in these products?
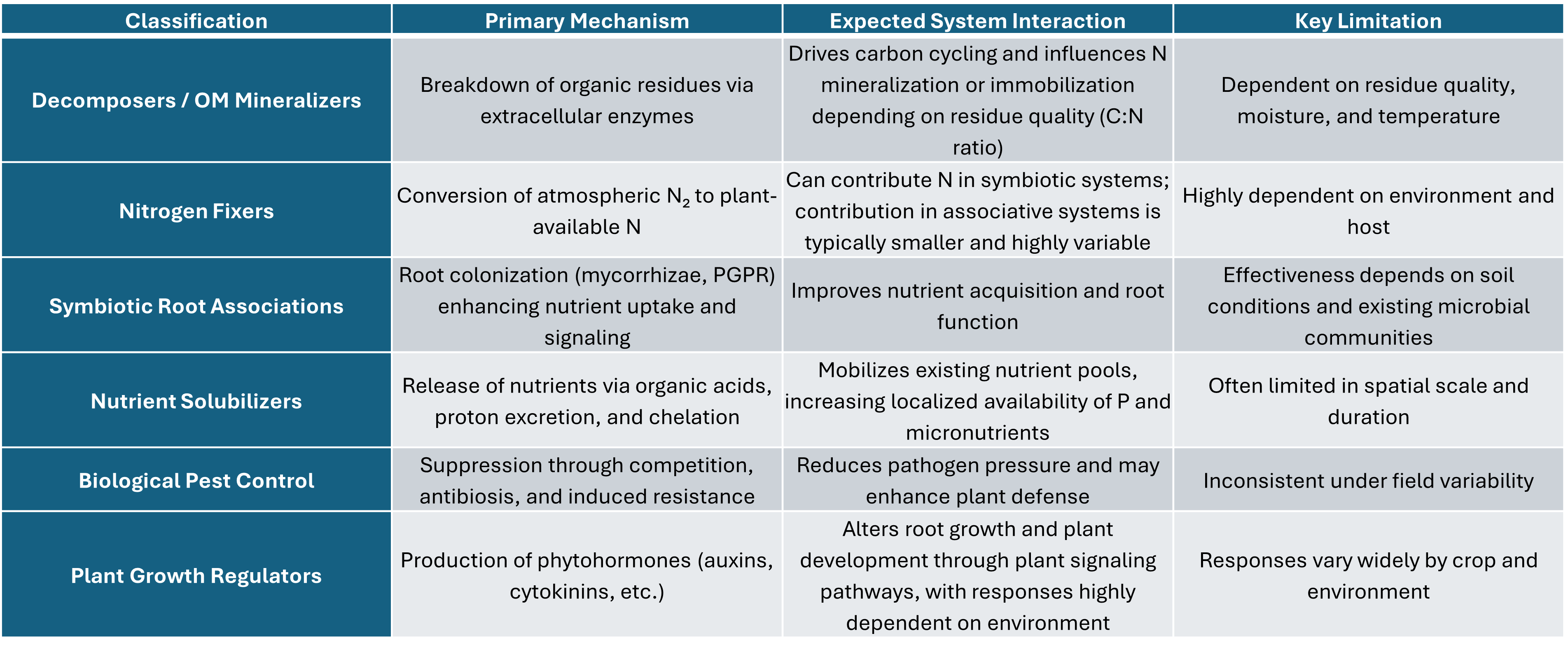
My approach has been to classify the products by operation not by species or genre. Doing so I have grouped products into five biological classifications and a sixth group, which is often in concluded in conversations.
Decomposers / Organic Matter Mineralizers
Nitrogen Fixers (Symbiotic and Associative)
Symbiotic Root Associations (Mycorrhizae, PGPR)
Nutrient Solubilizers
Biological Pest Control
Plant Growth Regulators (Hormonal Effects)
So, let’s dig into each of the mechanisms.
Decomposers / Organic Matter Mineralizers
Decomposition is carried out by a diverse group of organisms including fungi (e.g., Trichoderma, Aspergillus), bacteria (e.g., Bacillus, Pseudomonas), and actinomycetes (e.g., Streptomyces), each contributing to the breakdown of organic materials through different enzymatic pathways. This process of decomposing organic matter releases the nutrients tied up into plant available forms. The release of nitrogen is usually first thought, but this process adds significant amounts of potassium, calcium, and magnesium.
The process occurs both in the soil and on the soil surface. While it seems simple in application though this is a complex process. Let’s start with the soil pool, triggering decomposition of a system where the previous crop was wheat is significantly different than following corn. Following wheat, the carbon nitrogen ratio will be very high (see sugar blog), so while decomposition will release cations such as potassium and calcium, it is very likely to immobilize and residual nitrogen in the system. However, in fields that previously had corn the carbon to nitrogen ratio is much closer and the probability of seeing nitrogen release is much higher (Kuzyakov & Blagodatskaya, 2015). The process is similar for surface residues, but the rate is heavily controlled by rainfall. While both the soil and surface systems require moisture for the process to progress, the surface moisture is much more dynamic with frequent wetting and drying. Rain or irrigation is also needed to move the nutrients into the root zone.
One aspect of increasing decomposition of OM that I do not have a handle on is the long-term impact of expediting OM breakdown in and on the soil, especially in the central plains. As mentioned in the sugar blog, you would hope that the increase in nutrients from OM decomposition would increase plant growth enough to replenish the OM that was burned up. One caveat to this is that the decomposition would have to add nutrients that are deficient. Otherwise, there is no increase in plant growth and hypothetically the system is not net negative on OM. When it comes to decomposing surface residue, I have always been a bit hesitant in Oklahoma as I see having surface coverage to preserve soil moisture typically has a greater value than the nutrients from the residue.
Nitrogen Fixers (Symbiotic and Associative)
Nitrogen fixation is carried out by both symbiotic organisms such as Rhizobium and Bradyrhizobium, which form nodules on plant roots and supply significant nitrogen, and associative organisms such as Azospirillum and Azotobacter, which reside in the rhizosphere and contribute smaller, more variable amounts of nitrogen. Symbiotic nitrogen fixation, such as we have come to expect from legumes, is tightly regulated by the plant, with carbon supplied to the microbe in exchange for fixed nitrogen, making it one of the most efficient biological nitrogen inputs in agriculture.
Associative nitrogen fixation is not directly coupled to plant demand, and nitrogen contributions are typically limited by carbon availability and environmental conditions (Kennedy et al., 2004). While these organisms possess the ability to fix atmospheric nitrogen, the magnitude of nitrogen contribution, particularly from non-symbiotic systems, is highly variable and often limited under field conditions. We know that in soybean nodulation is greatly reduced when excess nitrogen is present in the soil, basically the plant does not need rhizobia, so it does not trigger symbiosis. I expect that as we move symbiotic fixation out of legumes that this mechanism does not change. Finally fixed N is no different than fertilizer N, if you add more then the crop needs, its lost. Therefore, if I am planning to use a N fixer, I would significantly reduce the amount of fertilizer N apply well below crop demand. Otherwise, the money spent on the N fixer is a waste. The only argument I have heard for this is the security blanket, making sure that if more is needed than normally the system is covered. But I circle back to the question about a system with high levels of residual N and rhizobium nodulation.
Symbiotic Root Associations (Mycorrhizae, PGPR)
Symbiotic root associations include arbuscular mycorrhizal fungi (e.g., Rhizophagus, Funneliformis) that extend the effective root system and improve nutrient uptake, particularly phosphorus, as well as plant growth-promoting rhizobacteria (e.g., Pseudomonas, Bacillus) that influence root development and plant signaling through multiple biochemical pathways (Smith & Read, 2008). In my visits with soil microbiologist, I have been left with the understanding that these relationships are not generic, but quite specific. There is significant influence of genotype and environment. And even more interesting is that the majority expect that the plant needs to signal for this relationship to happen.
The effectiveness of these associations is highly dependent on soil conditions, existing microbial communities, and nutrient availability, with responses often diminishing in systems where nutrients are not limited or where native populations are already established. I was able to follow along with some work down at OSU a few years back that was working with sorghum looking for symbiotic relationships to improve water and nutrient uptake specifically phosphorus. The work was successful, the researchers were able to identify a AMF that created a symbiotic relationship with sorghum, with a few caveats. First land race cultivars had a much higher incidence of symbiosis. For the landraces it worked well in extremely nutrient depleted soils and any additions of N or P reduced forage yield over the none. In the end the researchers were able to show improved the grain yield in landraces above fertilized, but these yields did equal fertilized hybrids. This work had great impact on small holders in developing counties with limited resources.
Nutrient Solubilizers
Nutrient solubilization is carried out by organisms such as Bacillus, Pseudomonas, and Aspergillus, which increase nutrient availability through mechanisms including organic acid production, proton release, and chelation, allowing nutrients like phosphorus and micronutrients to become more accessible in the rhizosphere.
Phosphorus-solubilizing fungi, such as Aspergillus and Penicillium, function similarly to bacterial solubilizers but are often more effective at producing strong organic acids. These acids can lower pH in localized zones and release phosphorus from mineral-bound forms, particularly in soils with high fixation capacity. Fungal systems can operate across a wider range of environmental conditions and may play a larger role in longer-term phosphorus cycling. However, as with bacterial systems, these effects are generally localized and dependent on soil chemistry (Richardson et al., 2009). I tend to see these having the greatest benefits in systems that have historically received manures or long-term applications of fertilizer P. I do not believe this is a good fit for soils with limited available phosphorus, as it is trying to focus the soil into something, it does not want to do or have too spare.
Potassium-solubilizing organisms, including species such as Bacillus mucilaginosus and Frateuria aurantia, contribute to the release of potassium from primary minerals like feldspars and micas. These microbes facilitate mineral weathering through acidification and chelation processes that slowly break down mineral structures. While the mechanism is well understood, the rate of potassium release is typically slow relative to crop demand. As a result, these organisms are more influential in long-term soil development than in short-term fertility management (Sheng & He, 2006).
Micronutrient-mobilizing organisms, particularly Pseudomonas and Bacillus species, enhance availability through the production of siderophores and other chelating compounds. These molecules bind metals such as iron and zinc, increasing their solubility and facilitating uptake in the rhizosphere. This process is especially important in soils where micronutrients are present but not readily available due to chemical constraints. However, the impact is typically limited to the immediate root zone and depends on both microbial activity and soil conditions (Ahmed & Holmström, 2014).
Biological Pest Control
Biological pest control organisms, including species such as Bacillus, Pseudomonas, and Trichoderma, function by suppressing pathogens through several well-documented mechanisms. These include the production of inhibitory compounds, competition for space and nutrients, direct antagonism of pathogens, and the activation of plant defense systems through induced systemic resistance. While these mechanisms are well established under controlled conditions, their effectiveness in field environments is highly dependent on environmental conditions, pathogen pressure, and the ability of the organism to persist and colonize the soil or plant surface (Lugtenberg & Kamilova, 2009).
I’ve been working with a lot of folks from Brazil who historically make four to six nemacide applications in soybean, but utilizing Pseudomanas they have been able to reduce that number by half or more. The caveat, as I understand, the application rates needed are significantly higher than anything I have seen in the US. If you look through the literature, you are seeing more and more documentation of this such as Li et al. 2022. But as Spescha et al. (2023) documented, different biological control agents operate through complementary mechanisms, including infection, toxin production, and host targeting. However, effectiveness depended on environmental conditions and interactions among organisms, reinforcing that biological control outcomes are system-dependent rather than universally consistent.
Plant Growth Regulators (Hormonal Effects)
This group differs slightly, as the primary effect is not direct nutrient cycling but modification of plant physiological response. This group is one I hold the greatest expectations for. I mean we have been using PGRs in crop production for decades, we just did not have an inkling of how many PGRs exist.
Plant growth regulator effects are associated with organisms such as Azospirillum, Bacillus, and Pseudomonas, which can influence plant development through the production of phytohormones and related compounds. These microbes produce substances such as auxins, cytokinins, and gibberellins that alter root architecture and plant growth patterns, and in some cases reduce stress responses through enzymes like ACC deaminase. Rather than supplying nutrients directly, these organisms modify how plants respond to their environment and utilize available resources. However, just like everything previously discussed the magnitude of response is often subtle and highly dependent on environmental conditions and crop system interactions (Glick, 2012).
Final thoughts.
There is one situation that pops up that I do not agree with, based upon my limited understanding of soil microbiology. Its adding more of what is already there. The soil system is a dynamic system. While there are population booms and bust, it supports what it is able to. Adding more of what is already there is like dropping a million rabbits into a prairie that has rabbits already. The current population is where it is because that is what the system can support. Adding means one of two things, a lot of rabbits die immediately, or they overwhelm the system and another animal species dies off due to lack of resources. Also, most microbiologists tell me the system is amazing at signaling and finding what it wants. It may take a season, but it will be there, in the quantities that soil needs, just given time.
So, the final slide in all my biological additives talks ends with this statement. My experiments show one thing. The impact of adding these products on crop yields is very consistently inconsistent. I’ve had many show a significant positive response, once. I have struggled to ever get repeated successes. It is my belief that I will have more success improving the soil biome by managing the soil (no-till, crop rotation, cover crops) than I will ever have with adding a product.
Final comment, Read the label. Many of the biological products I have tested are not singularly pure species. There are many blends of species and organisms which encompass many of the modes. A lot of these blends also contain extras such as humics, fulvics, carbohydrates, and sugars, see previous blogs.
Take-Home Messages
- Biological products function through specific mechanisms, not as broad “boosters,” and understanding that mechanism is critical to proper use.
- The presence of a biological function does not guarantee a yield response, outcomes are driven by soil, crop, and environmental conditions
- Decomposers and carbon-driven systems can immobilize or mineralize nitrogen, depending largely on residue quality and system balance
- Mycorrhizae and PGPR improve access to existing nutrients, not total nutrient supply
- Nutrient-solubilizing organisms mobilize nutrients already present in the soil
- Plant growth regulators influence plant signaling and development
- Adding biological organisms to soil does not guarantee establishment or persistence, as soil systems can regulate microbial populations.
- Management practices such as no-till, crop rotation, and cover crops are effective at improving soil biological function
- Across all biological products, mechanism exists, but response depends on the system
Any questions or comments please reachout to me @ b.arnall@okstate.edu
Citations
Ahmed, E., & Holmström, S. J. M. (2014). Siderophores in environmental research: Roles and applications. Microbial Biotechnology, 7(3), 196–208.
Glick, B. R. (2012). Plant growth-promoting bacteria: Mechanisms and applications. Scientifica, 2012, 963401
Kennedy, I. R., Choudhury, A. T. M. A., & Kecskés, M. L. (2004).
Non-symbiotic bacterial diazotrophs in crop-farming systems. Plant and Soil, 266, 65–79.
Kuzyakov, Y., & Blagodatskaya, E. (2015).
Microbial hotspots and hot moments in soil. Soil Biology and Biochemistry, 83, 184–199.
Lugtenberg, B., & Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annual Review of Microbiology, 63, 541–556.
Richardson, A. E., Barea, J. M., McNeill, A. M., & Prigent-Combaret, C. (2009). Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant and Soil, 321(1–2), 305–339.
Sheng, X. F., & He, L. Y. (2006). Solubilization of potassium-bearing minerals by a wild-type strain of Bacillus edaphicus and its mutants and increased potassium uptake by wheat. Canadian Journal of Microbiology, 52(1), 66–72. https://doi.org/10.1139/w05-117
Smith, S. E., & Read, D. J. (2008).
Mycorrhizal symbiosis. Academic Press.
Spescha, A., Weibel, J., Wyser, L., Brunner, M., Hess Hermida, M., Moix, A., Scheibler, F., Guyer, A., Campos-Herrera, R., Grabenweger, G., & Maurhofer, M. (2023). Combining entomopathogenic Pseudomonas bacteria, nematodes and fungi for biological control of a below-ground insect pest. Agriculture, Ecosystems & Environment, 348, 108414.
Ye S, Yan R, Li X, Lin Y, Yang Z, Ma Y and Ding Z (2022) Biocontrol potential of Pseudomonas rhodesiae GC-7 against the root-knot nematode Meloidogyne graminicola through both antagonistic effects and induced plant resistance. Front. Microbiol. 13:1025727. doi: 10.3389/fmicb.2022.1025727
The Mechanics of Soil Fertility: Use of Sugar in Field Crops
Jolee Derrick, Precision Nutrient Management Ph. D. Student
Grace Williams, Soil Microbiology Ph. D. Candidate
Brian Arnall, Precision Nutrient Management Specialist
Recently, there has been increased interest in adding sugar to spray tank mixes, whether for post-emergence weed control or foliar nutrient applications. While there is limited work on impact of sugar inclusion in herbicide applications, some papers have posed potential enhancement (Devine and Hall, 1990). But since this is coming from a soil science group, we will only focus on soil impact. Following up the last blog, unlike humic substances, which represent more complex and relatively stable carbon forms, sugar is a highly labile carbon source. This rapid utilization of simple carbon sources is well documented to stimulate microbial activity and growth (Kuzyakov and Blagodatskaya, 2015). The general idea of utilizing sugar applications is that sugar has the capacity to improve spray performance, stimulate biological activity, increase organic matter mineralization, and ultimately result in improved yields.
Sugar additions can influence soil processes differently depending on system conditions. In systems with higher residual nitrogen and organic matter, responses may differ from those observed in Oklahoma production environments, where soils are typically lower in organic matter and microbial activity can occur for much of the year. Understanding how sugar functions in these systems requires a basic discussion of carbon dynamics. Sugar itself is almost entirely carbon and is readily consumed by microbes. It’s a simple molecule, which allows it to dissolve easily in water and be quickly utilized in the soil system. Crop residues, like wheat straw, are also carbon-rich but much more complex. They contain cellulose, hemicellulose, and lignin which are long carbon chains that take time to break down because microbes need specialized enzymes to access them.
For the sake of simplicity, we can group carbon into two key pools: labile carbon and particulate organic matter (POM). Labile carbon includes easily decomposed materials, which include the previously mentioned simple sugars that microbes can metabolize rapidly. These pools differ in turnover time and microbial accessibility, with labile carbon driving short-term microbial responses (Cotrufo et al. 2013). POM breaks down more slowly and serves as a longer-term nitrogen source through residue breakdown.
Soil microorganisms require both carbon and nitrogen to grow and maintain biomass, typically at a ratio of approximately 24 parts carbon to 1 part nitrogen. When readily available carbon is abundant, but nitrogen is limited, microbes increase their nitrogen demand and begin scavenging nitrogen from the surrounding soil. This process, better known as nitrogen immobilization, temporarily reduces nitrogen availability to crops. Additions of readily available carbon sources have consistently been shown to increase microbial nitrogen immobilization in soil systems (Recous et al. 1990).
In systems where sufficient nitrogen is present, microbial populations can expand rapidly. Fast-growing microbial species may dominate, continuing to immobilize nitrogen within their biomass. Eventually, when nitrogen becomes limiting, microbial populations decline to levels the system can support. This boom-and-bust cycle can disrupt nitrogen availability during critical stages of crop growth. These rapid shifts in microbial population and activity following carbon inputs are commonly observed in soil systems receiving easily decomposable substrates (Blagodatskaya and Kuzyakov, 2008).
This dynamic becomes especially relevant when considering residue management practices common in Oklahoma. Under no-till or limited-tillage systems, the crop residues have wide carbon-to-nitrogen (C:N) ratios, creating conditions where nitrogen immobilization can occur during the growing season.
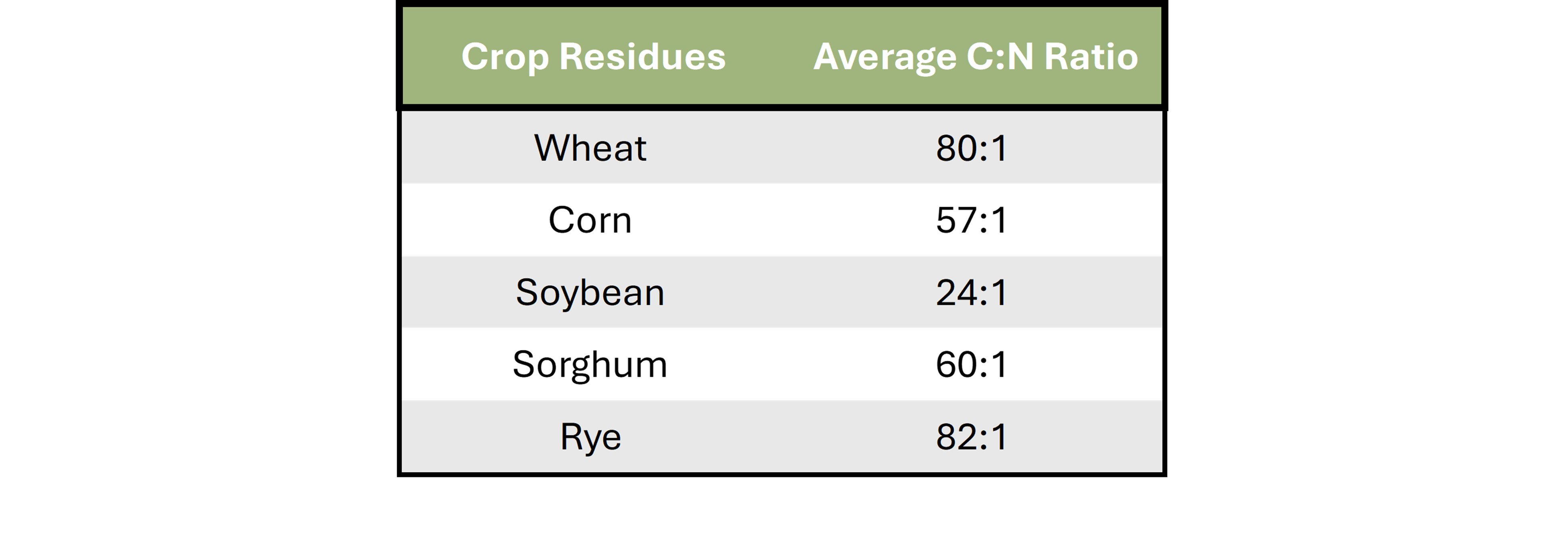
Table 1 provides approximate C:N ratios for several crops commonly grown in Oklahoma. When additional carbon is introduced into these systems without accompanying nitrogen, the likelihood of microbial immobilization increases. While immobilization is not bad, it does create a question mark as Oklahoma’s variable climate means the following release of nutrients will be unpredictable.
Table 1. Table depicting the range of C:N ratios for residues of commonly utilized crops in Oklahoma. Ratios were obtained from Brady, N. C., & Weil, R. R. (2017). The Nature and Properties of Soils (15th ed.)
Now consider conventional tillage systems. In Oklahoma, no-till systems typically contain 2 to 3 percent organic matter, which is relatively high given our climate and extended periods of microbial activity. Conventional tillage systems often fall between 0.75 and 2.25 percent organic matter. Because soil organic matter is approximately 58 percent carbon, this represents a substantial difference in the soil carbon pool.
Tillage can temporarily enhance microbial access to both previously mentioned carbon pools. When tillage exposes previously protected carbon, microbial activity increases rapidly. This initial flush can temporarily increase nitrogen mineralization as organic nitrogen is converted to plant-available forms. However, this phase is short-lived. As microbial populations expand, nitrogen demand increases, leading to immobilization and reduced nitrogen availability.
Hypothetically, increased microbial growth and activity would rapidly mineralize organic matter, trigger a surge in NO₃⁻, deplete soil organic matter, and as resources become limiting and the environment can no longer sustain elevated microbial populations, this boom would be followed by a population crash. This relationship is ultimately driven by the soil C:N ratio, which introduces an interesting additional complexity of residue. Different residues bring very different carbon-to-nitrogen balances into the system, and microbes respond accordingly. High carbon residues give microbes plenty of energy but very little nitrogen, so they pull N out of the soil to meet their needs. Residues with lower C:N ratios (soybean, alfalfa, etc.) do opposite, releasing nitrogen as they break down. Now the real question becomes where the critical point sits, and when does management push the system from the threshold of immobilization and mineralization.
These hypotheses form the foundation for new research currently underway through the Precision Nutrient Management Program. Initial proof-of-concept work has already been completed, providing a necessary steppingstone to address these questions.
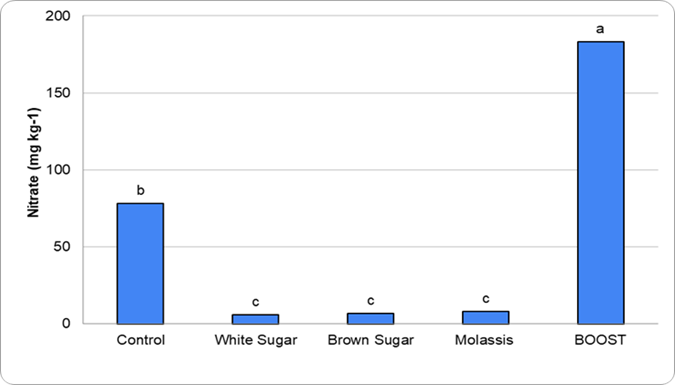
Figure 1. Graph depicting the different concentrations of nitrate leached corresponding to applied treatments in the proof-of-concept work
The preliminary work (Figure 1) evaluated different sugar sources applied alongside a high-nitrogen product to assess the extent of nitrogen immobilization. Although these studies were conducted using potting soils, clear trends were apparent. Treatments containing sugar consistently showed greater nitrogen immobilization compared to treatments without sugar. This response is consistent with studies showing that additions of simple carbon substrates stimulate microbial growth and increase nitrogen immobilization (Dendooven et al. 2006). Building on this work, an active field-based research project is underway to evaluate how sugar additions influence nitrogen availability and microbial dynamics under real-world Oklahoma production conditions.
From an agronomic standpoint, sugar functions primarily as a readily available carbon source that stimulates microbial growth. In nitrogen-limited systems, this response increases the likelihood that nitrogen will be incorporated into microbial biomass rather than remaining immediately available for crop uptake.
Finally, we conclude with a conceptual consideration. If increased OM mineralization leads to greater plant biomass, this process may partially offset losses of OM. Greater biomass production could return more residues to the soil, contributing to the OM pool in the upper soil profile. Therefore, the system may compensate for OM mineralization through the rebuilding of organic matter via plant inputs. However, the stabilization of this carbon depends on microbial processing and physical protection within the soil matrix (Cotrufo et al. 2015)
However, while the underlying logic is sound, this concept has not been extensively studied within Oklahoma cropping systems. This blog does not address the impact of sugar applications on residue breakdown, and the potential impact of such. Future research through the Precision Nutrient Management Program will further investigate the mineralization process to better understand carbon dynamics within these systems.
Take Home:
- Oklahoma production systems generally have lower residual N and high carbon residues, creating conditions conducive to N immobilization
- Adding sugar increases microbial growth, creating population booms that will momentarily increase mineralization, but then immediately immobilize residual nitrogen.
- Tillage can amplify the negative effects of sugar by exposing more carbon and reducing soil organic matter
- Proof-of-concept work shows sugar triggered a net nitrogen immobilization in a carbon heavy environment
- Proof-of-concept work also suggests that when additional nitrogen is present, sugar additions may shift the system toward net mineralization rather than immobilization.
Work Cited:
Blagodatskaya, E., & Kuzyakov, Y. (2008). Mechanisms of real and apparent priming effects. Biology and Fertility of Soils, 45, 115–131.
Brady, N. C., and R. R. Weil. “The Nature and Properties of Soils, 15th Edn (eBook).” (2017).
Cotrufo, M. F., Wallenstein, M. D., Boot, C. M., Denef, K., & Paul, E. (2013). The Microbial Efficiency-Matrix Stabilization (MEMS) framework. Global Change Biology, 19, 988–995.
Cotrufo, M. F., Soong, J. L., Horton, A. J., Campbell, E. E., Haddix, M. L., Wall, D. H., & Parton, W. J. (2015). Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nature Geoscience, 8(10), 776–779.
Dendooven, L., Verhulst, N., Luna-Guido, M., & Ceballos-Ramírez, J. M. (2006). Dynamics of inorganic nitrogen in nitrate- and glucose-amended alkaline–saline soil. Plant and Soil, 283(1–2), 321–333.
Devine, M. D., & Hall, L. M. (1990). Implications of sucrose transport mechanisms for the translocation of herbicides. Weed Science, 38(3), 299–304.
Kuzyakov, Y., & Blagodatskaya, E. (2015). Microbial hotspots and hot moments in soil: Concept & review. Soil Biology and Biochemistry, 83, 184–199.
Recous, S., Mary, B., & Faurie, G. (1990). Microbial immobilization of ammonium and nitrate in cultivated soils. Soil Biology and Biochemistry, 22, 913–922.
Monitoring for Cotton Jassid: A Potential New Threat to Oklahoma Cotton
Ashleigh M. Faris, Maxwell Smith, & Jenny Dudak
While the cotton jassid (Amrasca biguttula), also known as the Two-Spot Cotton Leafhopper, has not yet been detected in Oklahoma, its rapid expansion across the Southern Cotton Belt in 2025 makes it a potential threat to the 2026 season. This pest is considered one of the most serious threats to U.S. cotton, with the potential for significant yield losses in untreated fields. Stay informed on if the cotton jassid may be moving from the south and into Oklahoma by signing up for Texas A&M AgriLife Extension text alerts: text COTTON to 833.717.0325.
Identification: The “Two-Spot” Difference
The cotton jassid can be confused with the native potato leafhopper, but its unique markings are the key to early detection (Figure 1).
- Adults: Approximately 1/8 inch (2mm) long, wedge-shaped, and green.
- Key Markings: Look for two small black spots on the crown of the head and one black spot on the tip of each forewing. These spots on the head can sometimes fade but are generally visible under magnification; the spots on the wings will be present in adults and do not fade.
- Nymphs: Wingless and pale green. They are best known for their “crab-like” sideways movement when disturbed on the leaf surface. Any nymphs spotted should warrant a thorough scouting for adult cotton jassids and damage.

Figure 1. Cotton jassid adults (A) have one black dot on each wing and may have two small dots between their eyes (these dots on the crown can fade). The potato leafhopper (B), which is not a threat to cotton production but does occur in Oklahoma, does not have black dots on their wings or between their eyes. Image A courtesy of Isaac Esquivel, UF Extension, image B courtesy of DryBeanAgronomy.ca.
Biology and Host Range
The cotton jassid has a short life cycle, completing a generation in approximately two weeks under warm conditions.
- Reproduction: Eggs are inserted directly into leaf midveins and petioles, hatching in 3–4 days. Eggs will not be visible to the naked eye or through hand lens. Cotton jassids progress through 5 nymphal instars before becoming reproductive adults (Figure 2).
- Host Plants: This pest is polyphagous, meaning it feeds on many hosts. While cotton is a primary target, it also thrives on okra, eggplant, and ornamental hibiscus. It has also been found on native plants like Turk’s cap, as well as weeds like Ceasar weed and Florida pusley.
- 2025 Range on U.S. Cotton: The cotton jassid was detected on cotton in FL, GA, AL, MS, LA, TN, SC, NC, and TX. The TX detections in cotton were limited to southeastern TX in Grimes, Wharton, and Fort Bend counties. At the time of this article’s posting (March 2026), the cotton jassid has not been detected in OK.
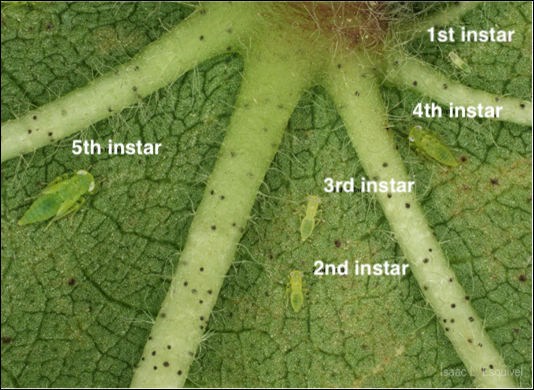
Figure 2. Cotton jassid nymphs on the underside of a cotton leaf. Image courtesy of Isaac Esquivel, UF Extension.
Damage: Recognizing Hopperburn
Unlike other leafhoppers, the cotton jassid injects a salivary toxin that disrupts the plant’s vascular system.
- Early Signs: Initial yellowing that resemble potassium deficiency with some upward curling of leaf margins (Figure 3, Rating 1).
- Progression: Characterized by hopperburn, a yellowing (chlorosis) that proceeds from leaf edges and turns red or brown as the tissue dies (Figure 3).
- Systemic Impact: Plants can go downhill quickly, often leading to complete desiccation and stunted growth (Figure 4).
A Hopperburn Injury Rating Scale has been developed by Extension Cotton Entomologists in the mid-South (Figure 3). You cannot let the cotton jassid get ahead of you. Once reddening starts on the leaf margins (Rating 2 in Figure 3) it is likely too late to rescue the cotton plant, damage will quickly progress and photosynthetic capabilities for the plant decline considerably.

Figure 3. Hopperburn injury rating scale for cotton jassid damage. Damage increases from none (0) to severe damage of desiccated leaf (5). Slight yellowing and upward curling of leaf is shown in Rating 1, with increased yellowing, cupping, and beginnings of reddened leaf margins in Rating 2. Insecticide action should be taken prior to reaching Rating 2. Ratings 3 – 5 show increased spread of reddening and desiccation. Images courtesy of Phillip Roberts (UGA Extension) and Scott Graham (AU Extension).

Figure 4. Cotton jassid hopperburn resulting in reddened, dried leaves (A) and stunted cotton plants (B). Image courtesy Isaac Esquivel, UF Extension.
Scouting Protocol
Scouting is mandatory for every cotton field in 2026 to prevent significant yield loss. Plants located at the edge of cotton fields can serve as good indicators, as cotton jassids will enter at field margins where damage is more likely to occur before further in field.
- Target Area: Inspect the undersides of leaves in the mid-to-upper canopy.
- Sample Leaf: Focus on the 4th mainstem leaf below the terminal, as this is where nymphs typically congregate.
- Visual Checks: Because adults fly quickly, count the flightless nymphs. Examine at least 25 leaves per field.
- Threshold: 1 cotton jassid per leaf, or early crop injury indicators (Figure 3, Rating 1) with cotton jassid confirmations nearby.
- Continue Scouting: Since green leaves are needed to fill bolls, growers should scout cotton up to at least 2 weeks prior to defoliation.
Management Guidance
Cultural Practices
- Plant Early: Trials indicate that earlier planting dates can help the crop “outrun” the peak pressure of migrating populations.
- Nutrient Management: Avoid excess Nitrogen, which attracts cotton jassids. Ensure adequate Potassium, as deficient plants crash much faster under cotton jassid stress.
- Varieties: Internationally, varieties with high trichome (hair) density on leaves offer natural resistance to feeding. However, varieties on the U.S. market are generally less hairy than those planted elsewhere. Currently, trials from 2025 do not indicate a varietal difference in terms of cotton jassid susceptibility.
Chemical Control
Based on 2025 research trials conducted by Mid-South Cotton Extension Entomologists, the following insecticides have shown varying levels of control (Table 1). Repeated insecticide applications may be warranted.
Table 1. Suggested foliar insecticides* and their observed control level for suppressing the cotton jassid. Efficacy lasted around 2 weeks.
| Control Level | Insecticides |
| High (>70% Control) | Carbine, Sefina, Sivanto, Bidrin, Venom, Plinazolin |
| Moderate (50-70%) | Transform, Centric, Assail, Orthene |
| Low (<50%) | Steward, Diamond, Bifenthrin, Admire Pro |
*Cotton jassids have shown resistance to every chemistry class in their native range; rotation of modes of action is critical. The mention, listing, or use of specific insecticides is not an endorsement of that product, nor is it a criticism of similar products not mentioned.
If you suspect cotton jassid activity or see hopperburn symptoms, contact the OSU Cotton IPM team: Maxwell Smith (maxwell.smith@okstate.edu), Ashleigh Faris (Ashleigh.faris@okstate.edu), and Jenny Dudak (jdudak@okstate.edu) immediately for confirmation. This team will be monitoring for the cotton jassid and will share updates on nearing threat, Oklahoma detections, and updated management guidance as it becomes available.
For more information on the cotton jassid in the U.S., click on this link to access Extension Factsheets, podcasts, and videos developed by Extension Entomologists managing the pest: https://drive.google.com/file/d/19IFT5c9b5JXEaBgf6X-weya07G56RXS5/view?usp=sharing.
Small Pest, Big Problems: Wheat Curl Mites and Wheat Streak Mosaic Virus Detected in Oklahoma
Ashleigh Faris, Cropping Systems Entomologist, IPM Coordinator
Meriem Aoun, Wheat Pathologist
Department of Entomology & Plant Pathology,
Oklahoma State University
Wheat Curl Mite (WCM) activity has been confirmed in Washita County, located in western Oklahoma. While the mites themselves are difficult to see, they can have a considerable impact on wheat health, primarily due to their role as vectors for several viral diseases such as wheat streak mosaic virus (WSMV). The Plant Disease and Insect Diagnostic Laboratory (PDIDL) has confirmed WSMV in the sample where WCM were detected in Washita County. This week, the PDIDL has also confirmed infection by WSMV in Blaine County (Canton, OK), McCurtain County (Garvin, OK), and Cleveland County (Noble, OK).
Identification
The Wheat Curl Mite is nearly invisible to the naked eye. At approximately 1/100 of an inch long, these pests require a 10x – 20x hand lens for proper identification.
- Appearance: They are white or cream-colored, cigar-shaped (cylindrical), and possess only four legs located near the head (Figure 1).
- Behavior: They are typically found in the protected areas of the plant, such as developing, youngest leaves or the furrows of the leaf surface. As the leaf unfurls, the mites migrate to the next emerging leaf.

Figure 1. Wheat curl mites and eggs on a wheat leaf (A, B), and mites on a maturing wheat kernel (C). Images courtesy G. Bauchan and R. Ochoa, USDA-ARS.
Biology and Life Cycle
Understanding the WCM life cycle is critical for preventative management:
- Rapid Reproduction: Under optimal temperatures (75° – 85°F), a WCM can complete its life cycle in 7 to 10 days. This allows populations to explode rapidly during warm autumns or springs.
- Dispersal: WCMs cannot fly; they rely entirely on wind currents to move from plant to plant or field to field. They crawl to the tips of leaves and hitchhike on the wind.
- Survival (The Green Bridge): WCMs are obligate parasites, meaning they require living green tissue to survive and reproduce. They persist through the summer on volunteer wheat and various perennial or annual grasses. This is known as the green bridge. If this bridge is not broken, mites move into the newly planted crop in the fall.
Damage and Virus Transmission
WCMs cause two types of damage:
- Direct Feeding: Mites suck sap from the leaf cells. This causes the edges of the leaf to roll inward (the curl part of WCM) (Figure 2). This curling provides a protected microclimate for the mites to reproduce. Heavy infestations can cause stunting and a slowed appearance in growth.
- Viral Vector (Primary Concern): The WCM is the sole vector for Wheat Streak Mosaic Virus (WSMV), High Plains Wheat Mosaic Virus (HPWMoV), and Triticum Mosaic Virus (TriMV).
- Symptoms: Infected plants show yellowing, mottled or streaked leaves, and severe stunting (Figures 3 & 4).
- Impact: If infection occurs in the fall, yield loss can be up to 100%. Spring infections are generally less damaging.
Scouting Techniques
Because the mites are so small, scouting focuses on leaf symptoms and having a hand lens:
- Check your Fields: Examine the youngest leaves of the wheat plant. Look for the characteristic inward rolling of the leaf edges (Figure 2).
- Use Magnification: Slowly unroll a suspect leaf and use a hand lens to look for tiny, white, slow-moving specks in the leaf furrows.
- Pattern of Infestation: Wind-dispersed mite infestations often start at the edge of a field (particularly edges adjacent to volunteer wheat or CRP land) and move inward in the direction of prevailing winds. Areas with infestations may show signs of yellowing and appear as patches distributed at random across the field (Figure 4).

Figure 2. Infestation of wheat curl mites on wheat results in tightly curled leaves and entrapment of subsequent leaves within the curl (A). After full leaf emergence, a tight curl at the leaf edge remains (B). Images courtesy of UNL Extension.

Figure 3. Wheat streak mosaic virus (WSMV) symptoms includeyellowing, mottled or streaked leaves. Image courtesy of Meriem Aoun, Oklahoma State University.

Figure 4. Plants at field margins, neighboring a wheat curl mite source, are the first to become infected with viruses of the Wheat Streak Mosaic Virus (WSMV) complex and develop symptoms, such as yellowing and streaking. Notice the gradient in color from the field edge (left) toward the center of the wheat field. Image courtesy of UNL Extension.
Management Recommendations
Currently, there are no effective rescue chemical treatments for WCM once symptoms appear in the field. Miticides generally do not reach the mites hidden inside the curled leaves. Management must be proactive:
- Manage volunteer wheat and grassy weeds: This is the most effective management tool to break the green bridge. Ensure all volunteer wheat and grassy weeds are completely dead (via tillage or herbicide) at least two weeks prior to planting the new crop. WCMs will starve within days without a living host.
- Delayed Planting: Planting wheat later in the fall reduces the window of time that mites must migrate into the crop and slows their reproduction rate as temperatures drop.
- Variety Selection: Some wheat varieties offer resistance or tolerance to WCM or WSMV. Consult the latest OSU variety trial data to select adapted varieties for north-central Oklahoma that carry these traits. Currently, Breakthrough is the most resistant OSU variety, which carries the WSMV resistance gene, Wsm1.
Brown Wheat Mite Activity in North Central Oklahoma
Ashleigh Faris, Cropping Systems Entomologist, IPM Coordinator
Department of Entomology & Plant Pathology,
Oklahoma State University
Following a period of dry weather, wheat growers in central Oklahoma are reporting activity of the Brown Wheat Mite (BWM). Unlike many other wheat pests, BWM thrives in drought conditions, and its damage can often be mistaken for moisture stress or nutrient deficiency.
Identification
The Brown Wheat Mite is small—about the size of a needle point—but is generally easier to spot than the Wheat Curl Mite because it is active on the leaf surface.
- Appearance: BWM has a dark red to brownish-black, oval-shaped body (Figure 1).
- Distinguishing Feature: Its front legs are significantly longer than its other three pairs of legs.
- Behavior: They are most active during the day, particularly in the afternoon, and will quickly drop to the ground if the plant is disturbed (Figure 2).

Figure 1. Brown wheat mite (BWM).

Figure 2. Brown wheat mites (BWM) on wheat. Image courtesy L. Galvin, OSU Extension.
Biology and Life Cycle
BWM populations consist entirely of females that produce offspring without mating (parthenogenesis), allowing for extremely rapid population growth under dry conditions. The BWM has a unique life cycle in that it can lay two types of eggs. Environmental conditions dictate when these two types of eggs are laid:
- Red Eggs: Laid during the growing season and hatch in about a week when conditions are favorable.
- White (Diapause) Eggs: Laid as temperatures rise and the crop matures. They are highly resistant and allow the population to survive the summer heat, hatching only when cooler, wetter weather arrives in the fall.
Damage
BWM damage is caused by the mites piercing plant cells and sucking out the plant nutrients.
- Symptoms: Initial damage appears as “stippling” (fine white or yellow spots) on the leaves. As feeding continues, leaves take on a silvery or bronzed appearance (Figure 3).
- Tipping: Heavy infestations cause the tips of the leaves to turn brown and die.
- Weather Interaction: Damage is most severe when plants are already under drought stress. Because both BWM damage and drought cause yellowing/browning, it is essential to confirm the presence of mites before treating.

Figure 3. Brown wheat mite (BWM) damage.
Scouting
Because BWM is highly mobile and drops when disturbed, careful scouting is required:
- Timing: Scout during the warmest part of the day when mites are most active on the upper leaves.
- The Paper Test: Gently but quickly shake or tap wheat plants over a white piece of paper or a white clipboard. Look for tiny dark specks moving across the surface.
- Economic Threshold: While thresholds vary based on crop value and moisture stress, research suggests a treatment threshold of 25 to 50 brown wheat mites per leaf in wheat that is 6 inches to 9 inches tall is economically warranted. An alternative estimation is “several hundred” per foot of row. If the wheat is severely stressed, the lower end of that threshold should be used.
Management Recommendations
- The “Rain” Factor: A significant, driving rain is often the most effective control for BWM. Rain can physically knock mites from the plant and promote fungal pathogens that naturally reduce the population.
- Chemical Control: If populations exceed the threshold and no rain is in the forecast, chemical intervention may be necessary. Know the cost of the treatment and value of your wheat so you can determine if an application is a worth return on investment.
- Effective Ingredients: Organophosphates (such as Dimethoate) have historically provided better control than many pyrethroids, as the latter can sometimes result in mite “flaring” or simply fail to provide adequate residual control.
- Coverage: High water volume is critical to ensure the insecticide reaches the mites, especially if they have moved toward the base of the plant.
- Pre-harvest Intervals & Grazing Restrictions: Always read and follow the label guidelines. For more on acaricides that can be applied in wheat see the Oklahoma State University Fact Sheet “Management of Insect and Mite Pests in Small Grains” (CR-7194).
- Cultural Practices: Since BWM thrives in dry, dusty conditions, maintaining good soil moisture and vigorous plant growth can help the crop tolerate feeding. Here’s to hoping for some rain soon in the forecast; we could really use it for lots of reasons in Oklahoma.
Check Your Wheat: Greenbugs Reported in Central Oklahoma
Ashleigh M. Faris
Cropping Systems Extension Entomologist
Department of Entomology & Plant Pathology
Oklahoma State University
Wheat producers in central Oklahoma are reporting the presence of the greenbug, Schizaphis graminum, in winter wheat fields. Greenbugs are one of the most important insect pests of wheat in the southern Great Plains and can occur from fall through spring. These aphids feed on plant sap and inject toxins into wheat plants, causing characteristic leaf discoloration and plant injury.
Early detection through field scouting is essential to determine whether populations are increasing and if an insecticide treatment is justified.
Greenbug Identification & Biology
Key identifying characteristics of greenbug (Figure 1):
- Small aphids (~1/16 inch long)
- Pale to lime-green body
- Dark green stripe down the middle of the back
- Dark tips on antennae and legs
- Found in colonies on the underside of wheat leaves

Greenbugs reproduce rapidly under favorable conditions (between 55° F and 95° F) and often occur in patches within fields rather than evenly distributed populations. During periods of cool weather, the greenbug may increase to enormous numbers, due to the absence of natural enemies, which develop significantly slower compared to greenbugs at such temperatures. On the other hand, cold weather can also influence aphid populations. However, this latest cold snap is not enough to eliminate greenbugs. It takes average temperatures below 20° F for at least a week to kill a substantial number of greenbugs in wheat.
Greenbug Damage in Wheat
Greenbugs damage wheat in two ways, through direct feeding and injection of toxic saliva. Greenbugs may also transmit barley yellow dwarf virus (BYDV), which can further reduce yield potential.
Typical early symptoms include small, reddish or copper spots on leaves (Figure 2) and yellowing around feeding sites. Advanced infestations will result in leaves turning yellow or orange, dead leaf tissue, stunted plants, and expanding patches of dead wheat. Heavy infestations may kill seedlings and reduce tillering, particularly during drought stress.

How to Scout for Greenbugs
The Glance-N-Go™ sampling system developed by Oklahoma State University can help determine whether aphid populations exceed economic thresholds. Download the Greenbug Glance N’ Go Sampler app for your smartphone. You will then input the control cost ($/Acre), crop value ($/Acre), and the Spring sampling window. Use a zig-zag or W-pattern (Figure 3) to scout your field, checking undersides of leaves at three tillers per stop for greenbugs and brown mummies. Use the app to record the numbers of these insects and sample until the app tells you to stop sampling or tells you treat. As temperatures warm, continue to scout regularly as greenbug populations may build.
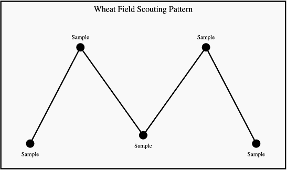
Scouting recommendations without the Greenbug Glance N’ Go Sampler app:
- Walk a W or zigzag pattern across the field.
- Examine 10–20 plants at each stop.
- Check:
- Underside of leaves
- Leaf midrib
- Base of tillers
- Record:
- Aphids per tiller
- Presence of aphid mummies (Figure 4)
- Beneficial insects
Beneficial Insects
Natural enemies frequently control aphid populations. While scouting for greenbug you should also look for lady beetles, lacewing larvae, hoverfly larvae, and parasitized aphids (“mummies”) (Figure 4). If beneficial insects are abundant, aphid populations may decline without insecticide treatment. Where there are one to two lady beetles (adults and larvae) per foot of row, or 15 to 20 percent of the greenbugs have been parasitized, control measures could be delayed until it is determined whether the greenbug population is continuing to increase.
Based on current wheat scouting, it appears that parasitoid numbers are low this 2026 season so continuing to scout for greenbug will be critical in responding to populations that go unchecked by beneficials.

Economic Threshold Guidelines
The simplest way to determine if action needs to be taken against greenbugs is to utilize the Glance-N-Go™ sampling system developed by Oklahoma State University. Approximate guidelines historically used in Oklahoma wheat can be found in Table 1 below.
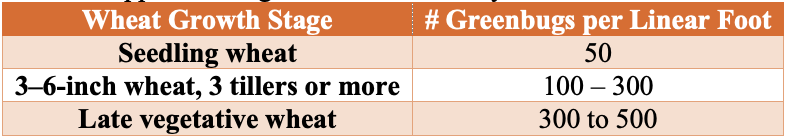
Thresholds are influenced by:
- Wheat growth stage
- Crop value
- Cost of treatment
- Presence of beneficial insects
Insecticides Labeled for Greenbugs in Wheat
Aphid feeding and insecticide performance are strongly influenced by temperature. Greenbugs tend to move higher on wheat plants during warm conditions but may move lower on the plant or below ground during cold weather, reducing exposure to insecticides. As a result, damaging populations are most often observed in late winter and early spring. Insecticides generally perform best when temperatures are above 50°F, and control may occur more slowly in cooler conditions (e.g., control at 45° F may take roughly twice as long as at 70° F). If applications must be made under cooler temperatures, use the highest labeled rate. Wheat grown under irrigation can typically tolerate higher greenbug populations than dryland wheat.
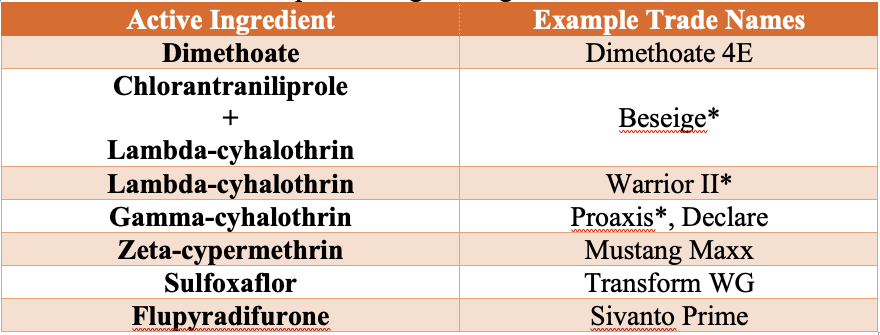
Always follow pesticide label directions, application sites, and rates. Be sure to read and follow the label for preharvest intervals (PHI) and restricted-entry intervals (REI). Use a minimum of 10 GPA by ground and 3 GPA by air (if labelled for aerial application) to ensure adequate coverage.
For assistance with aphid identification or treatment decisions, see OSU Fact Sheet EPP-7099 Small Grain Aphids in Oklahoma and Their Management, or contact your local OSU Extension office.
One Well-Timed Shot: Rethinking Split Nitrogen Applications in Wheat production
Brian Arnall, Precision Nutrient Management Specialist
Samson Abiola, PNM Ph.D. Student.
Nitrogen is the most yield limiting nutrient in wheat production, but it’s also the most unpredictable. Apply it too early, and you risk losing it to leaching or volatilization before your crop can use it. Apply it too late, and your wheat has already determined its yield potential; you’re just feeding protein at that point. For decades, the conventional wisdom has been to split nitrogen applications: put some down early to get the crop going, then come back later to apply again. But does splitting actually work? And more importantly, when is the optimal window to apply nitrogen if you want to maximize both yield and protein quality? We spent three years across different Oklahoma locations testing every timing scenario to answer these questions.
How We Tested Every Nitrogen Timing Scenario in Oklahoma Wheat
Between 2018 to 2021, we conducted field trials at three Oklahoma locations, including Perkins, Lake Carl Blackwell, and Chickasha, representing different soil types and growing conditions across the state. We tested three nitrogen rates: 0, 90, and 180 lbs N/ac, applied as urea at five critical growth stages based on growing degree days (GDD). These timings were 0 GDD (preplant, before green-up), 30 GDD (early tillering), 60 GDD (active tillering), 90 GDD (late tillering, approximately Feekes 5-6), and 120 GDD (stem elongation, approaching jointing). We also compared single applications at each timing against split applications, where half the nitrogen (45 lbs N ac-1) went down preplant, and the other half was applied in-season (45 lbs N ac-1).
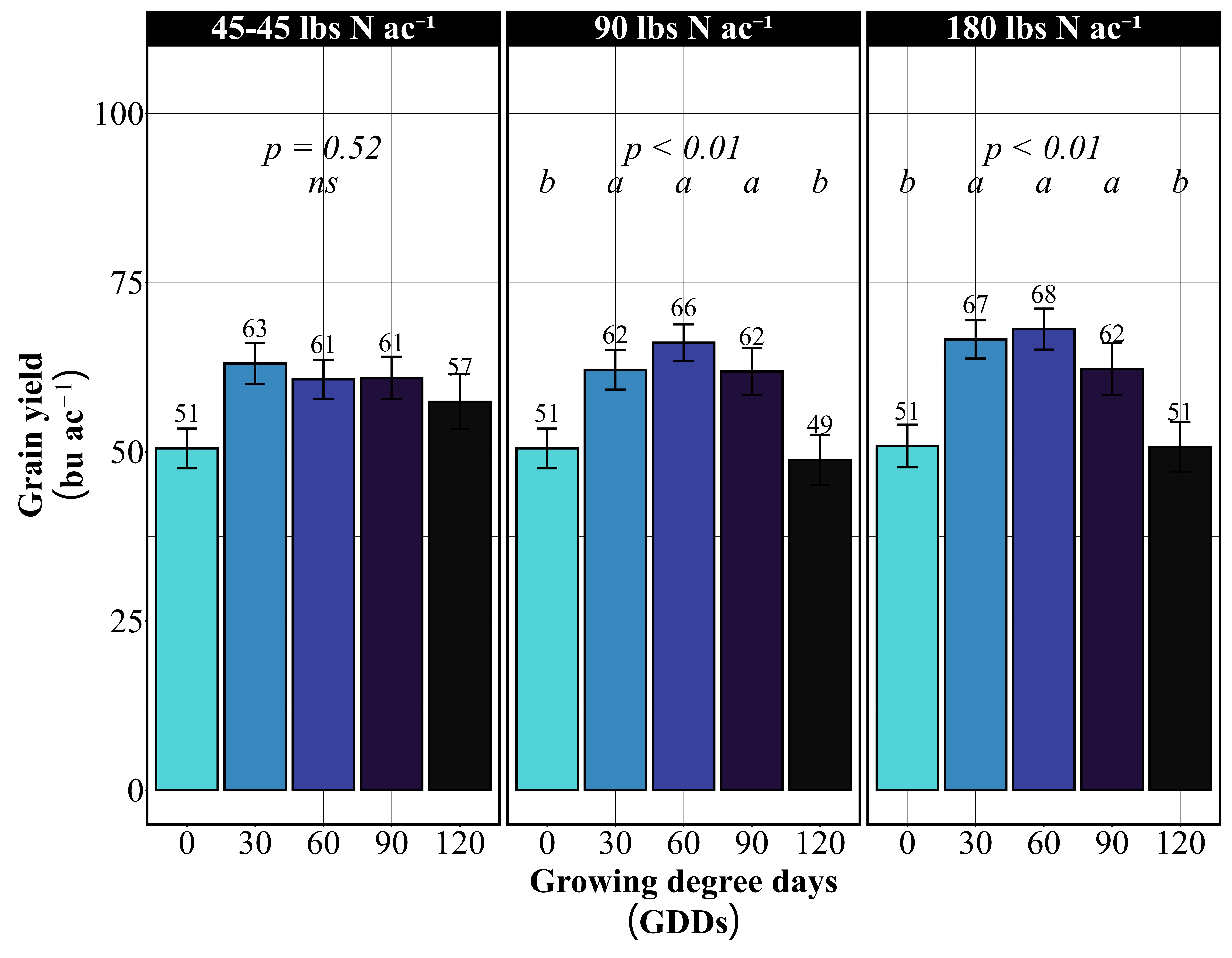
The Sweet Spot: Yield and Protein at the 90 lbs N/ac Rate
Across all site-years, at the 90 lbs N/ac rate, timing had a significant impact on both yield and protein. The highest yields came from the 30 and 90 GDD timings, producing 62 to 66 bu/ac, with 60 GDD reaching the peak (Figure 1). Protein at these early timings stayed relatively modest at 13%. The 90 GDD timing delivered 62 bu/ac with 14% protein matching the yield of the 30 GDD application but pushing protein a percentage higher (Figure 2). The real problem appeared at 120 GDD. Delaying application until stem elongation dropped yields to just 49 bu/ac, even though protein climbed to 15%. That’s a 13 bushel penalty compared to the 90 GDD timing. At current wheat prices per bushel, that late application may cost farmers over $100 per acre in lost revenue. By 120 GDD, the crop has already determined its yield potential tillers are set, head numbers are locked in and nitrogen applied at this stage can only be directed toward protein synthesis, not building more yield components.
More Nitrogen Does not lead to high yield
Doubling the nitrogen rate to 180 lbs N/ac revealed something critical, more nitrogen doesn’t mean more yield. The yield pattern remained nearly identical to the 90 lbs N/ac rate. The 60 GDD timing produced the highest yield at 68 bu/ac, followed closely by 30 GDD at 67 bu/ac. The 90 GDD timing yielded 62 bu/ac, and the 120 GDD timing again crashed to 51 bu/ac. The only difference between the two rates was protein concentration (Figure 2). At 180 lbs N/ac, protein levels increased across all timings: 13% at preplant, 15% at both 30 and 60 GDD, 15-16% at 90 GDD, and 16% at 120 GDD. This confirms a fundamental principle: once farmers supply enough nitrogen to maximize yield potential, which occurred at 90 lbs N/ac in these trials, additional nitrogen only increases grain protein. It does not build more bushels. Unless farmers are receiving premium payments for high-protein wheat, that extra 90 lbs of nitrogen represents a cost with no yield return.

Should farmers split their nitrogen application?
Now that timing has been established as critical, the next question becomes: should farmers split their nitrogen applications, or is a single application sufficient? The conventional recommendation has been to split nitrogen apply part preplant to support early growth and tillering, then return with a second application later in the season to boost protein and finish the crop. But does the data support this practice? We compared three strategies at each timing: applying all nitrogen preplant, applying all nitrogen in-season at the target timing, or splitting nitrogen equally between preplant and in-season timing. The goal was to determine whether the extra trip across the field will deliver better results.
Our findings revealed that splitting provided no consistent advantage. At 30 GDD, all three strategies preplant, in-season, and split performed identically, producing 62-65 bu/ac with 12-13% protein (Figure 3 and 4). No statistical differences existed among them. At 60 GDD, similar pattern was held. Yields ranged from 61 to 66 bu/ac and protein stayed at 12-13% regardless of whether farmers applied all nitrogen preplant, all at 60 GDD, or split between the two. At 90 GDD, the single in-season application actually outperformed the split. While yields remained similar across all three methods (61-64 bu/ac), the in-season application delivered significantly higher protein at 13.7% compared to 12.4% for preplant and 12.5% for split applications. This suggests that concentrating nitrogen at 90 GDD, rather than diluting it across two applications, allows more efficient incorporation into grain protein. The only timing where splits appeared beneficial was 120 GDD, where the split application yielded 59 bu/ac compared to 51 bu/ac for the single late application. But this is not a win for splitting, it simply demonstrates that applying all nitrogen at 120 GDD is too late and putting half down earlier salvages some of the yield loss. Across all timings tested, splitting nitrogen into two applications offered no agronomic advantage over a single well-timed application, meaning farmers are making an extra pass for no gain in yield or protein.


Practical Recommendations for Nitrogen Management
Based on three years of field data, farmers should target the 90 GDD timing (late tillering, Feekes 5-6) for their main nitrogen application to achieve the best balance between yield and protein. This window typically falls in late February to early March in Oklahoma, though farmers should monitor crop development rather than relying solely on the calendar apply when wheat shows multiple tillers, good green color, and vigorous growth. A rate of 90 lbs N/ac maximized yield in these trials; higher rates only increased protein without adding bushels, so farmers should only exceed this rate if receiving premium payments for high-protein wheat. Splitting nitrogen applications provided no advantage at any timing, meaning a single well-timed application at 90 GDD is sufficient for most Oklahoma wheat production systems. The exception would be sandy soils with high leaching potential, where splitting may reduce nitrogen loss. Farmers should avoid delaying applications until 120 GDD or later, as this timing consistently resulted in 15-25 bushel per acre yield losses even though protein increased. For farmers specifically targeting premium protein markets, a two-step strategy works best: apply 90 lbs N/ac at 90 GDD to establish yield potential and baseline protein, then follow with a foliar application of 20-30 lbs N/ac at flowering to push protein above 14% without sacrificing yield. Finally, weather conditions matter hot, dry forecasts increase volatilization risk and reduce uptake efficiency, so farmers should consider moving applications earlier if low humidity conditions are expected.
Split Application Caveat * Note from Arnall.
The caveat to the it only takes one pass, is high yielding >85+ bpa, environments. In these situation I still have not found any value for preplant nitrogen application. I have seen however a split spring application is valuable. Basically putting on 30-50 lbs at green-up, with the rest following at jointing (hollowstem). The method tends to reduce lodging in the high yielding environments.
This work was published in Front Plant Sci. 2025 Nov 6;16:1698494. doi: 10.3389/fpls.2025.1698494
Split nitrogen applications provide no benefit over a single well timed application in rainfed winter wheat
Another reason to N-Rich Strip.
Yet just one more data set showing the value of in-season nitrogen and why the N-Rich Strip concept works so well.
Questions or comments please feel free to reach out.
Brian Arnall b.arnall@okstate.edu
Acknowledgements:
Oklahoma Wheat Commission and Oklahoma Fertilizer Checkoff for Funding.
Double Crop Options After Wheat (KSU Edition)
Stolen from the KSU e-Update June 5th 2025.
Double cropping after wheat harvest can be a high-risk venture for grain crops. The remaining growing season is relatively short. Hot and/or dry conditions in July and August may cause problems with germination, emergence, seed set, or grain fill. Ample soil moisture this year can aid in establishing a successful crop after wheat harvest. Double-cropping forages after wheat works well even in drier regions of the state.
The most common double crop grain options are soybean, sorghum, and sunflower. Other possibilities include summer annual forages and specialized crops such as proso millet or other short-season summer crops, even corn. Cover crops are also an option for planting after wheat (see the companion eUpdate article “Cover crops grown post-wheat for forage”).
Be aware of herbicide carryover potential
One major planting consideration after wheat is the potential for herbicide carryover. Many herbicides applied to wheat are Group 2 herbicides in the sulfonylurea family with the potential to remain in the soil after harvest. If a herbicide such as chlorsulfuron (Glean, Finesse, others) or metsulfuron (Ally) has been used, then the most tolerant double crop will be sulfonylurea-resistant varieties of soybean (STS, SR, Bolt) or other crops. When choosing to use herbicide-resistant varieties, be sure to match the resistance trait with the specific herbicide (not only the herbicide group) that you used. This is especially true when looking at sunflowers as a double crop. There are sunflowers with the Clearfield trait, which allows Beyond herbicide applications, and ExpressSun sunflowers, which allow an application of Express herbicide. While both of these herbicides are Group 2 (ALS-inhibiting herbicides), the Clearfield trait and ExpressSun are not interchangeable, and plant damage can result from other Group 2 herbicides.
Less information is available regarding the herbicide carryover potential of wheat herbicides to cover crops. There is little or no mention of rotational restrictions for specific cover crops on the labels of most herbicides. However, this does not mean there are no restrictions. Generally, there will be a statement that indicates “no other crops” should be planted for a specified amount of time, or that a bioassay must be conducted prior to planting the crop.
Burndown of summer annual weeds present at planting is essential for successful double-cropping. Assuming glyphosate-resistant kochia and pigweeds are present, combinations of glyphosate with products such as saflufenacil (Sharpen) or tiafenacil (Reviton), or alternative treatments such as paraquat may be required. Dicamba or 2,4-D may also be considered if the soybean varieties with appropriate herbicide resistance traits are planted. In addition, residual herbicides for the double crop should be applied at this time.
Management, production costs, and yield outlooks for double crop options are discussed below.
Soybeans
Soybeans are likely the most commonly used crop for double cropping, especially in central and eastern Kansas (Figure 1). With glyphosate-resistant varieties, often the only production cost for planting double crop soybeans was the seed, an application of glyphosate, and the fuel and equipment costs associated with planting, spraying, and harvesting. However, the spread of herbicide-resistant weeds means additional herbicides will be required to achieve acceptable control and minimize the risk of further development of resistant weeds.

Weed control. The weed control cost cannot really be counted against the soybeans, since that cost should occur whether or not a soybean crop is present. In fact, having soybeans on the field may reduce herbicide costs compared to leaving the field fallow. Still, it is recommended to apply a pre-emergence residual herbicide before or at planting time. Later in the summer, a healthy soybean canopy may suppress weeds enough that a late-summer post-emergence application may not be needed.
Variety selection for double cropping is important. Soybeans flower in response to a combination of temperature and day length, so shifting to an earlier-maturing variety when planting late in a double crop situation will result in very short plants with pods that are close to the ground. Planting a variety with the same or perhaps even slightly later maturity rating (compared to soybeans planted at a typical planting date) will allow the plant to develop a larger canopy before flowering. Planting a variety that is too much later in maturity, however, increases the risk that the beans may not mature before frost, especially if long periods of drought slow growth. The goal is to maximize the length of the growing season of the crop, so prompt planting after wheat harvest time is critical. The earlier you can plant, the higher the yield potential of the crop if moisture is not a limiting factor.
Fertilizer considerations. Adding some nitrogen (N) to double-crop soybeans may be beneficial if the previous wheat yield was high and the soil N was depleted. A soil test before wheat harvest for N levels is recommended. Use no more than 30 lbs/acre of N. It would be ideal to knife-in the fertilizer. If that is not possible, banding it on the soil surface would be acceptable. Do not apply N in the furrow with soybean seed as severe stand loss can occur.
Seeding rates and row spacing. Seeding rate can be slightly increased if soybeans are planted too late in order to increase canopy development. Narrow row spacing (15-inch or less) has often resulted in a yield advantage compared to 30-inch rows in late plantings. Soybeans planted in narrow rows will canopy over more quickly than in wide rows, which is important when the length of the growing season is shortened. Narrow rows also offer the benefits of increasing early-season light capture, suppressing weeds, and reducing erosion. On the other hand, the advantage of planting in wide rows is that the bottom pods will usually be slightly higher off the soil surface to aid harvest. The other consideration is planting equipment. Often, no-till planters will handle wheat residue better and place seeds more precisely than drills, although the difference has narrowed in recent years.
What are typical yield expectations for double-crop soybeans? It varies considerably depending on moisture and temperature, but yields are usually several bushels less than full-season soybeans. A long-term average of 20 bushels per acre is often mentioned when discussing double-crop soybeans in central and northeast Kansas. Rainfall amount and distribution can cause a wide variation in yields from year to year. Double-crop soybean yields typically are much better as you move farther southeast in Kansas, often ranging from 20 to 40 bushels per acre.
A recent publication explores the potential yield of double-crop soybeans relative to full-season yield (Figure 2) and the most limiting factors affecting the yields for double-crop soybeans. The link to this article is: https://bookstore.ksre.ksu.edu/pubs/MF3461.pdf.

Grain Sorghum
Grain sorghum is another double crop option. Unlike soybeans, sorghum hybrids for double cropping should be earlier maturing hybrids. Sorghum development is primarily driven by the accumulation of heat units, and the double crop growing season is too short to allow medium-late or late hybrids to mature before the first frost in most of Kansas.
Seeding rates and row spacing. Late-planted sorghum likely will not tiller as much as early plantings and can benefit from slightly higher seeding rates than would be used for sorghum planted at an earlier date. Narrow row spacing is advised, especially if the outlook for rainfall is good.
Fertilizer considerations. A key component for the estimation of N application rates is the yield potential. This will largely determine the N needs. It is also important to consider potential residual N from the wheat crop. This can be particularly important when wheat yields are lower than expected. In that situation, additional available N may be present in the soil. Assess the amount of profile N by taking soil samples at a depth of 24 inches and submitting them for analysis at a soil testing laboratory.
Double crop sorghum planted into average or greater-than-average amounts of wheat residue can result in a challenging amount of residue to deal with when planting next year’s crop. Nitrogen fertilizer can be tied up by wheat residue, so use application methods to minimize tie-up, such as knifing into the soil below the residue.
Weed control. Weed control can be important in double-crop sorghum. Warm-season annual grasses, such as crabgrass, can reduce double-crop sorghum yields. Using a chloroacetamide-and-atrazine pre-emergence product may be key to successful double-crop sorghum production. Herbicide-resistant grain sorghum varieties will allow the use of imazamox (Imiflex in igrowth sorghums) or quizalofop (FirstAct in DoubleTeam grain sorghum) that can control summer annual grasses.
No-till studies at Hesston documented 4-year average double crop sorghum yields of 75 bushels per acre compared to about 90 bushels per acre for full-season sorghum. A different 10-year study that did not have double crop planting but did compare early- and late-planting dates averaged 73 bushels per acre for May planting vs. 68 bushels per acre for June planting.
Sunflowers
Sunflowers can be a successful double crop option anywhere in the state, provided there is enough moisture at planting time to get a stand. Sunflowers need more moisture than any other crop to germinate and emerge because of the large seed. Therefore, stand establishment is important. Planting immediately after wheat harvest on a limited irrigation field can be a good fit to help with stand establishment.
Seeding rates and hybrid selection. When double-cropping sunflowers, producers should use similar seeding rates to what is typical for the area for full-season sunflowers. While full-season sunflowers can be successful in double-crop production, utilizing shorter-season hybrids can increase the likelihood of the sunflowers blooming and maturing before a killing frost.
Weed control. First, it is important to check the herbicide applications on the wheat. The rotation restriction to sunflowers after several commonly used wheat herbicides is 22-24 months.
Weed control can be an issue with double-crop sunflowers since herbicide options are limited, especially post-emergence. Thus, controlling weeds prior to sunflower planting is critical and may be complicated pre-plant restrictions for some herbicides. Planting Clearfield or ExpressSun sunflowers will provide additional post-emergence herbicide options, but ALS-resistant kochia and pigweeds still won’t be controlled. Imazamox (Beyond in Clearfield sunflower) has activity on small annual grasses as well as many broadleaf weeds, if they are not ALS-resistant.
Summer annual forages
With mid-July plantings, and where herbicide carryover issues are not a concern, summer annual sorghum-type forages are also a good double crop option. A study planted July 21, 2008 near Holton, when summer rainfall was very favorable, provided yields of 2.5 to 3 tons dry matter/acre for hybrid pearl millet and sudangrass at the low end to 4 to 5 tons dry matter/acre for forage sorghum, BMR forage sorghum, photoperiod sensitive forage sorghum, and sorghum x sudangrass hybrids. Earlier plantings may produce even more tonnage, as long as there is adequate August rainfall.
One challenge with late-planted summer annual forages is getting them to dry down when harvest is delayed until mid- to late-September. Wrapping bales or bagging to make silage are good ways to deal with the higher moisture forage this late in the year.
Corn
Is double-crop corn a viable option? Corn is typically not recommended for late June or July plantings because yield is usually substantially less than when planted earlier.
Typically, mid-July planted corn struggles during pollination and seldom receives sufficient heat units to fill grain before frost. Very short-season corn hybrids (80 to 95 RM) have the greatest chance of maturing before frost in double crop plantings, but generally have less yield potential when compared to hybrids of 100 RM or more used for full-season plantings. Short-season hybrids often set the ear fairly close to the ground, increasing the harvest difficulty. Glyphosate-resistant hybrids will make weed control easier with double crop corn, but problems remain present with late-emerging summer weeds such as pigweeds, velvetleaf, and large crabgrass. Keep in mind, corn is very susceptible to carryover of most residual ALS herbicides used in wheat.
Considerations for altering seeding rates and variety/hybrid maturity for the crops discussed above are summarized in Table 1.
Table 1. Seeding rate and variety/hybrid relative maturity considerations for double crops compared to full-season.
| Crop | Seeding rate | Relative maturity |
| ???????? Difference between double crop and full-season ???????? | ||
| Soybean | Increase | No change or longer |
| Sorghum | Increase | Shorter |
| Sunflower | No change | Shorter |
| Corn | No change | Shorter |
Volunteer wheat control
One of the issues with double cropping that is often overlooked by producers is the potential for volunteer wheat in the crop following wheat. If volunteer wheat emerges and goes uncontrolled, it can cause serious problems for nearby wheat fields in the fall as a host for the wheat streak mosaic complex of viruses [wheat streak mosaic (WSMV), High Plains disease (HPD), and triticum mosaic (TriMV)] that are transmitted by the wheat curl mite (WCM).
Volunteer wheat can generally be controlled fairly well with glyphosate or Group 1 herbicides such as quizalofop (Assure II, others), clethodim (Select Max, others), or sethodydim (Poast Plus, others), but control is reduced during times of drought stress. Atrazine can provide control of volunteer wheat in double-crop corn or sorghum, but control can be erratic depending on rainfall patterns.
For more detailed information about herbicides, see the “2025 Chemical Weed Control for Field Crops, Pastures, and Noncropland” guide available online at https://www.bookstore.ksre.ksu.edu/pubs/CHEMWEEDGUIDE.pdf or check with your local K-State Research and Extension office for a paper copy. The use of trade names is for clarity to readers and does not imply endorsement of a particular product, nor does exclusion imply non-approval. Always consult the herbicide label for the most current use requirements.
To Subscribe to the KSU Agronomy E-Updates follow this link
https://eupdate.agronomy.ksu.edu/index_new_prep.php
Authors contributing to the post
Sarah Lancaster, Weed Management Specialist
slancaster@ksu.edu
John Holman, Cropping Systems Agronomist
jholman@ksu.edu
Logan Simon, Southwest Area Agronomist
lsimon@ksu.edu
Tina Sullivan, Northeast Area Agronomist
tsullivan@ksu.edu
Jeanne Falk Jones, Multi-County Agronomist
jfalkjones@ksu.edu
