Brian Arnall, Oklahoma State University Precision Nutrient Management
The use of biological products in commercial agriculture has expanded rapidly, with large corporations entering a space once dominated by smaller groups. This has created an arms race, with nearly every company offering a biological product. Over the past twenty years, I have had the opportunity to test products from the biggest groups with billions in backing, to solutions raised in stock tanks delivered in Braums milk jugs. It is critical to understand what is in the jug and the biological function it is expected to perform. Like herbicides, knowing the mode of action determines whether the product fits the intended purpose. No different than herbicides and knowing mode of actions. It’s important to know and understand that if you are trying to kill ryegrass 2.4-D, a broadleaf herbicide is not the right answer.
So what are we working with that’s in these products?
My approach has been to classify the products by operation not by species or genre. Doing so I have grouped products into five biological classifications and a sixth group, which is often in concluded in conversations.
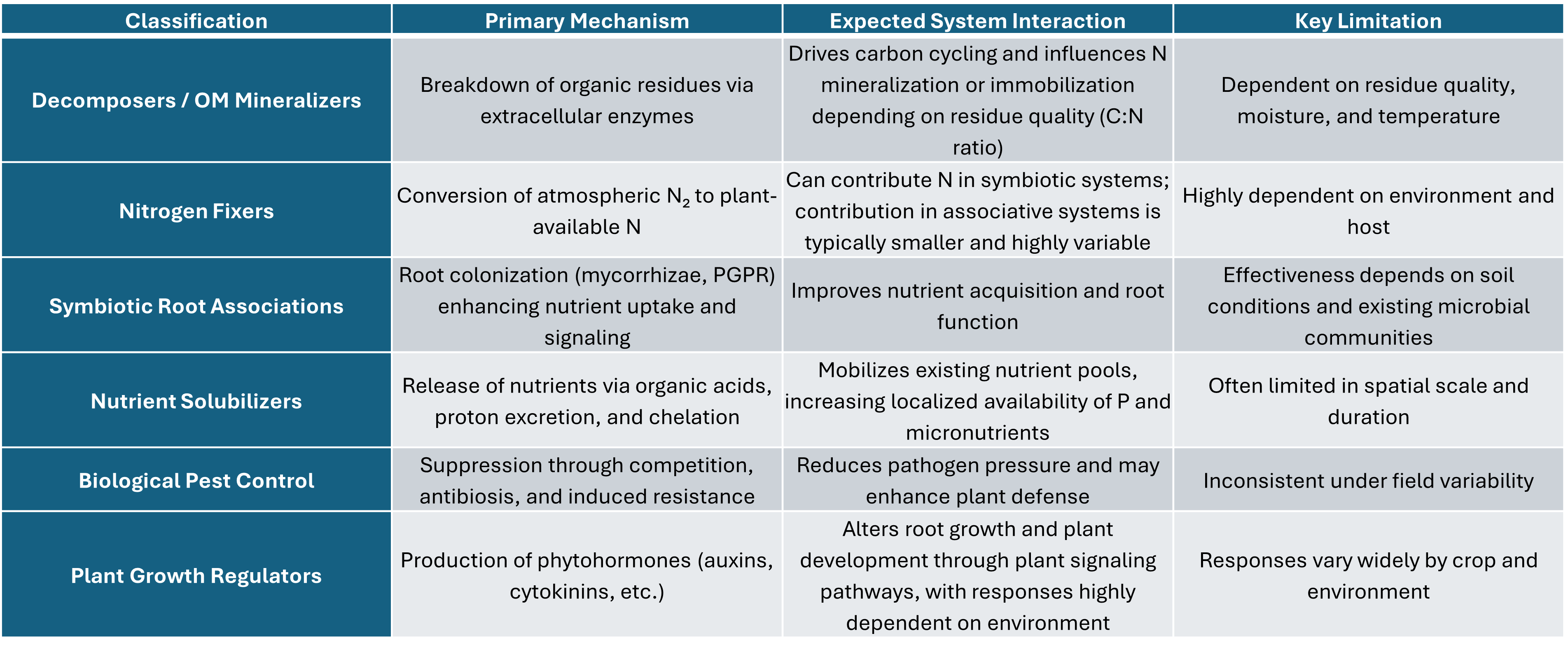
Decomposers / Organic Matter Mineralizers
Nitrogen Fixers (Symbiotic and Associative)
Symbiotic Root Associations (Mycorrhizae, PGPR)
Nutrient Solubilizers
Biological Pest Control
Plant Growth Regulators (Hormonal Effects)
So, let’s dig into each of the mechanisms.
Decomposers / Organic Matter Mineralizers
Decomposition is carried out by a diverse group of organisms including fungi (e.g., Trichoderma, Aspergillus), bacteria (e.g., Bacillus, Pseudomonas), and actinomycetes (e.g., Streptomyces), each contributing to the breakdown of organic materials through different enzymatic pathways. This process of decomposing organic matter releases the nutrients tied up into plant available forms. The release of nitrogen is usually first thought, but this process adds significant amounts of potassium, calcium, and magnesium.
The process occurs both in the soil and on the soil surface. While it seems simple in application though this is a complex process. Let’s start with the soil pool, triggering decomposition of a system where the previous crop was wheat is significantly different than following corn. Following wheat, the carbon nitrogen ratio will be very high (see sugar blog), so while decomposition will release cations such as potassium and calcium, it is very likely to immobilize and residual nitrogen in the system. However, in fields that previously had corn the carbon to nitrogen ratio is much closer and the probability of seeing nitrogen release is much higher (Kuzyakov & Blagodatskaya, 2015). The process is similar for surface residues, but the rate is heavily controlled by rainfall. While both the soil and surface systems require moisture for the process to progress, the surface moisture is much more dynamic with frequent wetting and drying. Rain or irrigation is also needed to move the nutrients into the root zone.
One aspect of increasing decomposition of OM that I do not have a handle on is the long-term impact of expediting OM breakdown in and on the soil, especially in the central plains. As mentioned in the sugar blog, you would hope that the increase in nutrients from OM decomposition would increase plant growth enough to replenish the OM that was burned up. One caveat to this is that the decomposition would have to add nutrients that are deficient. Otherwise, there is no increase in plant growth and hypothetically the system is not net negative on OM. When it comes to decomposing surface residue, I have always been a bit hesitant in Oklahoma as I see having surface coverage to preserve soil moisture typically has a greater value than the nutrients from the residue.
Nitrogen Fixers (Symbiotic and Associative)
Nitrogen fixation is carried out by both symbiotic organisms such as Rhizobium and Bradyrhizobium, which form nodules on plant roots and supply significant nitrogen, and associative organisms such as Azospirillum and Azotobacter, which reside in the rhizosphere and contribute smaller, more variable amounts of nitrogen. Symbiotic nitrogen fixation, such as we have come to expect from legumes, is tightly regulated by the plant, with carbon supplied to the microbe in exchange for fixed nitrogen, making it one of the most efficient biological nitrogen inputs in agriculture.
Associative nitrogen fixation is not directly coupled to plant demand, and nitrogen contributions are typically limited by carbon availability and environmental conditions (Kennedy et al., 2004). While these organisms possess the ability to fix atmospheric nitrogen, the magnitude of nitrogen contribution, particularly from non-symbiotic systems, is highly variable and often limited under field conditions. We know that in soybean nodulation is greatly reduced when excess nitrogen is present in the soil, basically the plant does not need rhizobia, so it does not trigger symbiosis. I expect that as we move symbiotic fixation out of legumes that this mechanism does not change. Finally fixed N is no different than fertilizer N, if you add more then the crop needs, its lost. Therefore, if I am planning to use a N fixer, I would significantly reduce the amount of fertilizer N apply well below crop demand. Otherwise, the money spent on the N fixer is a waste. The only argument I have heard for this is the security blanket, making sure that if more is needed than normally the system is covered. But I circle back to the question about a system with high levels of residual N and rhizobium nodulation.
Symbiotic Root Associations (Mycorrhizae, PGPR)
Symbiotic root associations include arbuscular mycorrhizal fungi (e.g., Rhizophagus, Funneliformis) that extend the effective root system and improve nutrient uptake, particularly phosphorus, as well as plant growth-promoting rhizobacteria (e.g., Pseudomonas, Bacillus) that influence root development and plant signaling through multiple biochemical pathways (Smith & Read, 2008). In my visits with soil microbiologist, I have been left with the understanding that these relationships are not generic, but quite specific. There is significant influence of genotype and environment. And even more interesting is that the majority expect that the plant needs to signal for this relationship to happen.
The effectiveness of these associations is highly dependent on soil conditions, existing microbial communities, and nutrient availability, with responses often diminishing in systems where nutrients are not limited or where native populations are already established. I was able to follow along with some work down at OSU a few years back that was working with sorghum looking for symbiotic relationships to improve water and nutrient uptake specifically phosphorus. The work was successful, the researchers were able to identify a AMF that created a symbiotic relationship with sorghum, with a few caveats. First land race cultivars had a much higher incidence of symbiosis. For the landraces it worked well in extremely nutrient depleted soils and any additions of N or P reduced forage yield over the none. In the end the researchers were able to show improved the grain yield in landraces above fertilized, but these yields did equal fertilized hybrids. This work had great impact on small holders in developing counties with limited resources.
Nutrient Solubilizers
Nutrient solubilization is carried out by organisms such as Bacillus, Pseudomonas, and Aspergillus, which increase nutrient availability through mechanisms including organic acid production, proton release, and chelation, allowing nutrients like phosphorus and micronutrients to become more accessible in the rhizosphere.
Phosphorus-solubilizing fungi, such as Aspergillus and Penicillium, function similarly to bacterial solubilizers but are often more effective at producing strong organic acids. These acids can lower pH in localized zones and release phosphorus from mineral-bound forms, particularly in soils with high fixation capacity. Fungal systems can operate across a wider range of environmental conditions and may play a larger role in longer-term phosphorus cycling. However, as with bacterial systems, these effects are generally localized and dependent on soil chemistry (Richardson et al., 2009). I tend to see these having the greatest benefits in systems that have historically received manures or long-term applications of fertilizer P. I do not believe this is a good fit for soils with limited available phosphorus, as it is trying to focus the soil into something, it does not want to do or have too spare.
Potassium-solubilizing organisms, including species such as Bacillus mucilaginosus and Frateuria aurantia, contribute to the release of potassium from primary minerals like feldspars and micas. These microbes facilitate mineral weathering through acidification and chelation processes that slowly break down mineral structures. While the mechanism is well understood, the rate of potassium release is typically slow relative to crop demand. As a result, these organisms are more influential in long-term soil development than in short-term fertility management (Sheng & He, 2006).
Micronutrient-mobilizing organisms, particularly Pseudomonas and Bacillus species, enhance availability through the production of siderophores and other chelating compounds. These molecules bind metals such as iron and zinc, increasing their solubility and facilitating uptake in the rhizosphere. This process is especially important in soils where micronutrients are present but not readily available due to chemical constraints. However, the impact is typically limited to the immediate root zone and depends on both microbial activity and soil conditions (Ahmed & Holmström, 2014).
Biological Pest Control
Biological pest control organisms, including species such as Bacillus, Pseudomonas, and Trichoderma, function by suppressing pathogens through several well-documented mechanisms. These include the production of inhibitory compounds, competition for space and nutrients, direct antagonism of pathogens, and the activation of plant defense systems through induced systemic resistance. While these mechanisms are well established under controlled conditions, their effectiveness in field environments is highly dependent on environmental conditions, pathogen pressure, and the ability of the organism to persist and colonize the soil or plant surface (Lugtenberg & Kamilova, 2009).
I’ve been working with a lot of folks from Brazil who historically make four to six nemacide applications in soybean, but utilizing Pseudomanas they have been able to reduce that number by half or more. The caveat, as I understand, the application rates needed are significantly higher than anything I have seen in the US. If you look through the literature, you are seeing more and more documentation of this such as Li et al. 2022. But as Spescha et al. (2023) documented, different biological control agents operate through complementary mechanisms, including infection, toxin production, and host targeting. However, effectiveness depended on environmental conditions and interactions among organisms, reinforcing that biological control outcomes are system-dependent rather than universally consistent.
Plant Growth Regulators (Hormonal Effects)
This group differs slightly, as the primary effect is not direct nutrient cycling but modification of plant physiological response. This group is one I hold the greatest expectations for. I mean we have been using PGRs in crop production for decades, we just did not have an inkling of how many PGRs exist.
Plant growth regulator effects are associated with organisms such as Azospirillum, Bacillus, and Pseudomonas, which can influence plant development through the production of phytohormones and related compounds. These microbes produce substances such as auxins, cytokinins, and gibberellins that alter root architecture and plant growth patterns, and in some cases reduce stress responses through enzymes like ACC deaminase. Rather than supplying nutrients directly, these organisms modify how plants respond to their environment and utilize available resources. However, just like everything previously discussed the magnitude of response is often subtle and highly dependent on environmental conditions and crop system interactions (Glick, 2012).
Final thoughts.
There is one situation that pops up that I do not agree with, based upon my limited understanding of soil microbiology. Its adding more of what is already there. The soil system is a dynamic system. While there are population booms and bust, it supports what it is able to. Adding more of what is already there is like dropping a million rabbits into a prairie that has rabbits already. The current population is where it is because that is what the system can support. Adding means one of two things, a lot of rabbits die immediately, or they overwhelm the system and another animal species dies off due to lack of resources. Also, most microbiologists tell me the system is amazing at signaling and finding what it wants. It may take a season, but it will be there, in the quantities that soil needs, just given time.
So, the final slide in all my biological additives talks ends with this statement. My experiments show one thing. The impact of adding these products on crop yields is very consistently inconsistent. I’ve had many show a significant positive response, once. I have struggled to ever get repeated successes. It is my belief that I will have more success improving the soil biome by managing the soil (no-till, crop rotation, cover crops) than I will ever have with adding a product.
Final comment, Read the label. Many of the biological products I have tested are not singularly pure species. There are many blends of species and organisms which encompass many of the modes. A lot of these blends also contain extras such as humics, fulvics, carbohydrates, and sugars, see previous blogs.
Take-Home Messages
- Biological products function through specific mechanisms, not as broad “boosters,” and understanding that mechanism is critical to proper use.
- The presence of a biological function does not guarantee a yield response, outcomes are driven by soil, crop, and environmental conditions
- Decomposers and carbon-driven systems can immobilize or mineralize nitrogen, depending largely on residue quality and system balance
- Mycorrhizae and PGPR improve access to existing nutrients, not total nutrient supply
- Nutrient-solubilizing organisms mobilize nutrients already present in the soil
- Plant growth regulators influence plant signaling and development
- Adding biological organisms to soil does not guarantee establishment or persistence, as soil systems can regulate microbial populations.
- Management practices such as no-till, crop rotation, and cover crops are effective at improving soil biological function
- Across all biological products, mechanism exists, but response depends on the system
Any questions or comments please reachout to me @ b.arnall@okstate.edu
Citations
Ahmed, E., & Holmström, S. J. M. (2014). Siderophores in environmental research: Roles and applications. Microbial Biotechnology, 7(3), 196–208.
Glick, B. R. (2012). Plant growth-promoting bacteria: Mechanisms and applications. Scientifica, 2012, 963401
Kennedy, I. R., Choudhury, A. T. M. A., & Kecskés, M. L. (2004).
Non-symbiotic bacterial diazotrophs in crop-farming systems. Plant and Soil, 266, 65–79.
Kuzyakov, Y., & Blagodatskaya, E. (2015).
Microbial hotspots and hot moments in soil. Soil Biology and Biochemistry, 83, 184–199.
Lugtenberg, B., & Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annual Review of Microbiology, 63, 541–556.
Richardson, A. E., Barea, J. M., McNeill, A. M., & Prigent-Combaret, C. (2009). Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant and Soil, 321(1–2), 305–339.
Sheng, X. F., & He, L. Y. (2006). Solubilization of potassium-bearing minerals by a wild-type strain of Bacillus edaphicus and its mutants and increased potassium uptake by wheat. Canadian Journal of Microbiology, 52(1), 66–72. https://doi.org/10.1139/w05-117
Smith, S. E., & Read, D. J. (2008).
Mycorrhizal symbiosis. Academic Press.
Spescha, A., Weibel, J., Wyser, L., Brunner, M., Hess Hermida, M., Moix, A., Scheibler, F., Guyer, A., Campos-Herrera, R., Grabenweger, G., & Maurhofer, M. (2023). Combining entomopathogenic Pseudomonas bacteria, nematodes and fungi for biological control of a below-ground insect pest. Agriculture, Ecosystems & Environment, 348, 108414.
Ye S, Yan R, Li X, Lin Y, Yang Z, Ma Y and Ding Z (2022) Biocontrol potential of Pseudomonas rhodesiae GC-7 against the root-knot nematode Meloidogyne graminicola through both antagonistic effects and induced plant resistance. Front. Microbiol. 13:1025727. doi: 10.3389/fmicb.2022.1025727
